





Polymorphism
Polymorphism is very common among cryptic animals. It is almost universal in grasshoppers, mantises, and stick insects. It is found in land snails, marine crabs, leaf-hoppers, spiders, lizards, frogs, and mice (reviewed in Bond 2007, Diamond & Bond 2013). What these creatures have in common is that they are often preyed upon by animals, such as birds, fishes, and reptiles, that tend to hunt by sight. Edward Poulton (1890) first noted this pattern and suggested that it could be a general evolutionary response to visual predation. He reasoned that it was "harder and more time-consuming" for a bird to search for several different targets simultaneously than to search for only one, so that it would be an advantage for prey species to occur in multiple forms. Luuk Tinbergen (1960) directly observed patterns of predation by insect-eating birds and inferred that the birds were confining their search to only one or two prey types at any one time. To maximize their rate of detection, the birds focused on the most common prey available (or the one that they had encountered most recently) and effectively overlooked the others, a process Tinbergen called "hunting by searching image."
One hypothesis we have tested extensively in our laboratory is that hunting by searching image should tend to promote polymorphism in previously uniform populations, because it will select against individuals that bear a close resemblance to one another. This evolutionary mechanism has been termed "apostatic selection," because it tends to overlook individuals with a unique appearance (i.e. "apostates"; Clarke 1962). Because predators tend to focus their search on the most conspicuous and most abundant forms, while allowing more cryptic and rarer forms to survive and continue breeding, apostatic selection will not necessarily result in extinction of any of the prey morphs. Instead, the population should stabilize around a specific set of morphs arrayed in order of how difficult they are to detect -- cryptic forms will be more abundant, but even relatively conspicuous forms should still survive at some low level. The result should be "balanced polymorphism," but until we ran experiments on a realistic search task with polymorphic prey, there was no direct evidence of its existence.
To study the effects of searching image on the dynamics of a prey species, we created a virtual population of 200 digital moths (Bond & Kamil 1998). Our initial population contained equal numbers of each of three distinctive moth types. As in our earlier work, the moths were presented singly on cryptic backgrounds to blue jays in an operant chamber. To obtain a realistic "virtual ecology," however, four different blue jays were used as predators, with each bird seeing a randomly-selected quarter of the moth population each day. Detected individuals were considered "killed" and were removed from the population. Moths that were overlooked were allowed to breed, in proportion to the relative abundance of their moth type, bringing the population up to its previous level the following day. Each day thus constituted a generation. Our primary experimental intervention was to set the initial numbers of the three morphs.
 |
| Moth images used in the apostatic selection experiment. Moths 1, 2, and 3 constituted the initial prey population. Moths 4 and 5 were novel forms that were introduced in later stages of the experiment. Note that Moth 5 is considerably different in appearance from the other four. |
This methodology defines a fixed phenotype or "coexistence" procedure (Kassen 2002), in which the population of digital moths consists of a set of asexually reproducing clones of invariant appearance. They do not mutate, and each generation is brought up to a constant size based on the relative numbers of surviving individuals. The population dynamics of the different morphs and their asymptotic levels of abundance were, thus, the main dependent variables. Over the course of 50 generations, the numbers of the three morphs rapidly achieved a characteristic equilibrium that was independent of initial relative abundances and resistant to subsequent perturbation.
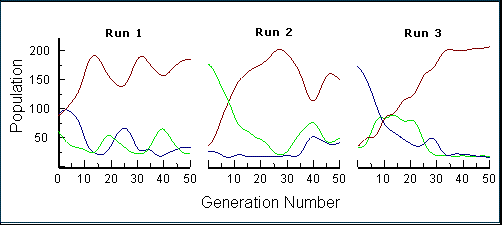 |
| In the first 50-generation run, three moth types were initially set to equal abundances. Within 10-15 generations, they had shifted to characteristic equilibrium levels. In subsequent runs, one of the less abundant moths at equilibrium was initially set high to perturb the system. In each case, the previous equilibrium levels were rapidly regained, demonstrating stable system dynamics. |
Additional analyses demonstrated that the equilibrium was in fact a result of apostatic selection -- a negative feedback between prey abundance and detectability. If jays tend to concentrate their searching efforts on more common moth types, we would expect that the proportion of individuals of a specified type that were discovered (="taken") by the birds would be higher than expected for common forms and lower than expected for rarer ones. So there should be a significant linear relationship between the deviation from randomness ("proportion taken minus proportion present") and the proportion present in the population. This relationship was confirmed for all three moth types (see below), providing the first direct demonstration of the dynamic relationship between searching image, apostatic selection, and polymorphic stability.
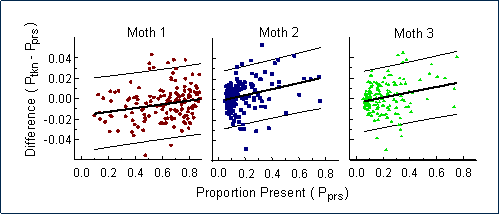 |
We then tested the effects of apostatic selection on novel morphs, i.e. moth types that the birds had not seen previously. (Bond & Kamil 1998). When small numbers of each a new moth were introduced into the population, they were not initially detected by the jays and their abundance rapidly increased. In the first case (Moth 4), the jays ultimately took notice of the new prey items and drove their numbers down, establishing a new equilibrium state. The second novel form we tried had a different outcome, however. Moth 5 rapidly increased until it completely dominated the population. Even after 100 generations, only two of the four jays had learned to find it, and if we had not continuously reintroduced small numbers of the original three forms, they would all have been driven to extinction (see below).
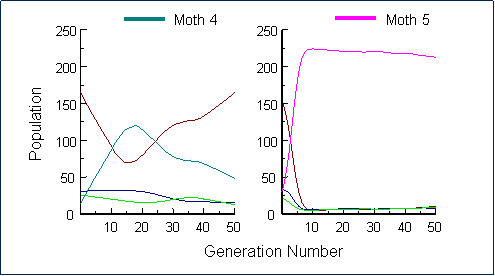 |
From these results, the stability of the observed configuration of morphs was clearly a function of their familiarity to the birds, or at least of their resemblance to known, familiar forms. Rare, novel, inconspicuous morphs may take a long time to be discovered by predators and may, in consequence, disrupt a previous stable polymorphism. Whether this would prove to be the case in a more realistic system in which prey phenotypes were free to evolve was not clear. To explore this question, we needed to extend our procedures to enable the evolution of prey appearance, and for that, we needed to develop a virtual genetics.
References from Other Sources
Clarke, B. (1962). Balanced polymorphism and the diversity of sympatric species. In D. Nichols, ed., Taxonomy and Geography, 47-70. Oxford: Systematics Association Publications.
Kassen, R. (2002). The experimental evolution of specialists, generalists, and the maintenance of diversity. Journal of Evolutionary Biology 15: 173-190.
Poulton, E.B. (1890). The Colours of Animals: Their Meaning and Use, Especially Considered in the Case of Insects. New York: Appleton.
Tinbergen, L. (1960). The natural control of insects in pine woods I. Factors influencing the intensity of predation by songbirds. Arch. Néerl. Zool. 13: 265343.