|
||
| Content Pages | ||
|
Social Cognition Social Inference Symbolic Inference |
||
|
|
||
| People | ||
|
Alan B. Bond Alan C. Kamil Russell P. Balda Jeffrey R. Stevens |
||
|
|
||
| Docs & Links | ||
|
Books Publications Links |
||
|
|
||
 |
||
Symbolic Transitive Inference
In Bond, Kamil and Balda (2003), we tested pinyon jays and scrub jays on an abstract version of a dominance hierarchy, an implicit ordinal sequence of seven colored stimuli. The sequence was implicit, in that birds were only shown two stimuli at a time. During training, only adjacent pairs were presented. The sequence was ordered, in that birds were only rewarded if they chose to peck the higher member of the pair (Fersen et al. 1991; Treichler & Van Tilburg 1996). Thus, on one trial, the bird could be shown a blue and a red circle, and he would be rewarded for pecking the blue one. On another trial, he would see a red and a green circle, and in this case he would be rewarded for pecking the red one. Different color sequences were used for each bird, to control for possible differences in how easily particular colors could be distinguished.
The birds were given 36 pair-comparison trials per day. They were trained initially on the A/B pair until they reached sufficient levels of accuracy. Subsequently, they were given the B/C pair, and so on through the six successive dyads in the hierarchy. When they were fully trained on all adjacent pairs, we trained them in sessions with mixtures of two adjacent pairs (e.g. A/B and B/C). Although it is difficult to compare the results of these training trials to other studies, scrub jays appeared to learn the color relationships somewhat faster than pigeons (Fersen et al. 1991), while pinyon jays were only slightly slower than rhesus monkeys (Treichler & Van Tilburg 1996). At the start of the third phase of the experiment, the two species were equally accurate and well above chance levels.
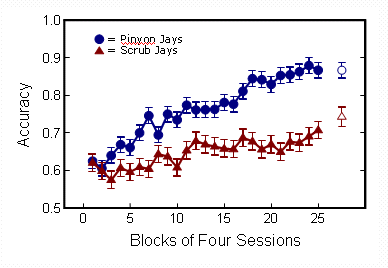
In the third phase, the birds were given 100 sessions in which all six color pairs were presented equally often. Pinyon jays improved more rapidly than scrub jays and showed a higher over-all level of performance. In the graph below, the filled symbols show the average accuracy across birds for each species over this phase of the experiment. The open figures show the results of an additional 100 sessions, indicating that the species difference was still present even after over a year of training on the seven stimuli. This result is consistent with the first prediction of the social complexity hypothesis: that highly social animals should be able to keep accurate track of a larger number of dyadic relationships.
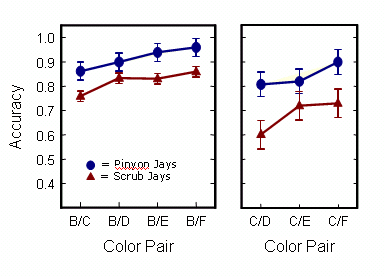
We then turn to the second prediction, which concerns transitive inference. The criterion of transitive reasoning here is fairly simple: After training only on adjacent dyads, animals should be able to respond correctly to novel stimulus pairings, pairs that they had never previously seen, thereby displaying a knowledge of the underlying ordinal sequence. Previous work on other species suggests that transitive inferences are generally stronger and more accurate for stimuli that are further apart on the implicit sequence, what has been called the Symbolic Distance effect (Moyer & Bayer 1976), These data are shown on the graph below. In the left panel, the X axis shows pairs of stimuli at progressively greater distances from one another along the implicit sequence, all beginning with the second-ranked stimulus. The right panel shows the same effect beginning with the third-ranked stimulus. Notice that both species show a clear increase in accuracy with increasing distance along the implicit sequence. Which is to say that both jays show some ability to make transitive inferences (Fersen et al. 1991; Bryant & Trabasso 1971; Steirn, Weaver & Zentall 1995).
This is not the end of the story, however, because there is evidence in the literature for two different cognitive mechanisms for transitive inference. An associative mechanism (Terrace & McGonigle 1994) simply works on the basis of frequency of reward, how often responses to the given stimulus have been rewarded. By any of several models of this cognitive mechanism, the net result should be a gradient of associative strength that is higher at the beginning of the sequence and drops off toward the end (Fersen et al. 1991; Zentall & Sherburne 1998; Delius & Siemann 1998). More importantly, however, the variance in associative strength is higher toward the end of the sequence. And this means that an associative representation should become less accurate the further down the sequence you go, what has been called a First Item accuracy effect (Fersen et al. 1991). This has been shown several times in pigeons (Fersen et al. 1991; Steirn, Weaver & Zentall 1995; Wynne 1997; Delius & Siemann 1998).
In contrast, monkeys commonly seem to insert the results of paired contrasts into an underlying linear array, so that the relationships between the stimuli are directly encoded and the stimuli are associated with their ordinal positions. This has been termed a linear (D’Amato & Columbo 1990) or relational (D’Amato 1991) mechanism. Monkeys show no obvious first item accuracy effects, but apparently because they find the relative positions of two stimuli by counting down from the beginning of the sequence, they often show a First Item effect on response time (D’Amato & Columbo 1988, 1990; D’Amato 1991). So we have a straightforward method of distinguishing the two cognitive mechanisms, based on the effect of the position of the highest member of the stimulus pair.
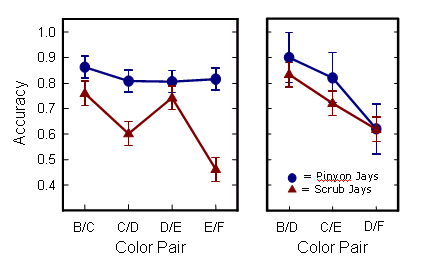
The First Item results for accuracy are plotted below. Scrub jays show a significant first item accuracy effect, both for adjacent pairs (on the left) and non-adjacent pairs (on the right). Pinyon jays show no significant effects of first item ranking on accuracy. The apparent trend for pinyons in the right panel is due to a single bird and is not statistically significant. Scrub jays thus appear to use the pigeon-like mechanism of associative representation.
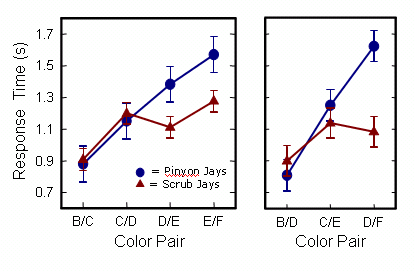
And below are the First Item results for correct response time, again with adjacent pairs on the left and non-adjacent pairs on the right. The two species do not differ in response time at the top of the sequence (B/C or B/D), but they progressively diverge, and toward the end of the sequence (D/E, E/F, or D/F), pinyon jays are taking significantly longer to make correct choices. Pinyon jays thus appear to be using a monkey-like relational or linear mechanism.
The conclusions from this study are that pinyon jays learn to track multiple dyadic relationships more rapidly and more accurately than scrub jays. And they appear to use a more robust and sophisticated mode of transitive inference, one that is less susceptible to error across the entire stimulus sequence. These results provide the first quantitative demonstration of a clear association between social complexity and cognitive representation in animals.
References from Other Sources:
Bryant, P.E. & Trabasso, T. 1971. Transitive inferences and memory in young children. Nature, 232, 456-458.
D’Amato, M.R. 1991. Comparative cognition: Processing of serial order and serial pattern. In: Current Topics in Animal Learning: Brain, Emotion, and Cognition (Ed. by L. Dachowski & C. F. Flaherty), pp. 165-185. Hillsdale, NJ: Erlbaum.
D’Amato, M.R. & Columbo, M. 1988. Representation of serial order in monkeys (Cebus apella). Journal of Experimental Psychology: Animal Behavior Processes, 14, 131-139.
D’Amato, M.R. & Columbo, M. 1990. The symbolic distance effect in monkeys (Cebus apella). Animal Learning & Behavior, 18, 133-140.
Delius, J.D. & Siemann, M. 1998. Transitive responding in animals and humans: Exaptation rather than adaptation? Behavioural Processes, 42, 107-137.
Fersen, L. von, Wynne, C.D.L., Delius, J.D. & Staddon, J.E.R. 1991. Transitive inference formation in pigeons. Journal of Experimental Psychology: Animal Behavior Processes, 17, 334-341.
Moyer, R.S. & Bayer, R.H. 1976. Mental comparison and the symbolic distance effect. Cognitive Psychology, 8, 228-246.
Steirn, J.N., Weaver, J.E. & Zentall, T.R. 1995. Transitive inference in pigeons: Simplified procedures and a test of value transfer theory. Animal Learning & Behavior, 23, 76-82.
Terrace, H.S. & McGonigle, B. 1994. Memory and representation of serial order by children, monkeys, and pigeons. Current Directions in Psychological Science, 3, 180-185.
Treichler, F.R. & Van Tilburg, D. 1996. Concurrent conditional discrimination tests of transitive inference by macaque monkeys: List linking. Journal of Experimental Psychology: Animal Behavior Processes, 22, 105-117.
Wynne, C.D.L. 1997. Pigeon transitive inference: Tests of simple accounts of a complex performance. Behavioural Processes, 39, 95-112.
Zentall, T.R. & Sherburne, L.M. 1998. The transfer of value in simultaneous discriminations: Implications for cognitive and social processes. In: Brain and Values: Is a Biological Science of Values Possible? (Ed. by K. H. Pribram), pp. 323-336. Hillsdale, NJ: Erlbaum.